


I. Introduction
The resurgence of the clinical use of perchlorate as a means of controlling amiodarone-induced thyroid dysfunction has suggested a need for a more careful examination of the actions of this antithyroid anion. To accomplish this, it is essential to understand the chemical, biochemical, and physiological bases for the action of perchlorate in the iodine economy of the thyroid gland. Many of its lateantithyroid effects leading to hypothyroidism will not be discussed as they differ little from those of the thionamides. On the other hand, the early effects in the pathway of iodine metabolism differ markedly from those of the thionamides, and it is these aspects that are the subject of this review. Perchlorate competitively interferes with iodide accumulation in the thyroid; as a result, it blocks hormone production and output like the more familiar thionamide drugs but by a different mechanism. This property has led to a deeper understanding of thyroid gland function, particularly the iodide transport system and is the basis for its clinical usefulness in treating thyrotoxicosis, particularly when induced by an iodide load. For the same mechanistic reason, perchlorate ion is used to discharge accumulated iodide from the thyroid that has iodide organification defects.
II. The Anion Transport System
To place the functions of perchlorate into proper perspective, a brief summary of the iodide concentrating mechanism is required, for it shares this mechanism with perchlorate. One of the crucial specializations of thyroid tissue is its ability to concentrate iodide selectively from the surroundings where the concentration is very low (10−8 – 10−7m) and where the concentration of chloride ions will be of the order of 0.01 to 0.1 m. The thyroid has thus developed a mechanism that can establish intrathyroidal iodide concentrations up to several hundred-fold those of the external milieu. Such concentrations are presumably required to promote efficient hormone synthesis (Wolff et al., 1959, 1972) because patients lacking the ability to concentrate iodide have goiters and are hypothyroid (reviewed in Wolff, 1983; a dozen or so additional cases have been reported since Alberto et al., 1987; Couch et al., 1985; Léger et al., 1987; Vulsma et al., 1991; Wémeau et al., 1991). This genetic defect can be entirely corrected by providing large doses of iodide that enter the gland by diffusion. The ability to concentrate iodide is shared by some other tissues, such as salivary glands, gastric mucosa, and the choroid plexus (Brown-Grant, 1961), some of which have readily accessible secretions that can be sampled for diagnostic purposes (Wolff, 1983). Nevertheless, it is essentially only in the thyroid gland that the newly concentrated iodide can be further metabolized to form thyroid hormone. This selective transport system can be conveniently divided into two mechanistically distinct properties: those aspects dealing with energetics and those dealing with specificity for anions i.e., recognition. The latter is more directly pertinent to the perchlorate anion. For recent reviews see Ekholm, 1990; Taurog, 1996; Wolff, 1992; Wollman, 1995.
A. Energetics
Thyroidal iodide transport is “active” not only by electrochemical criteria (Chow et al., 1969; Wolff, 1964; Wright and Diamond, 1977), but also by numerous metabolic criteria: it does not occur in the cold, requires O2 (Surks, 1967), and is a function of the adenosine triphosphate (ATP)b level (Wolff and Halmi, 1963; Mayaan and Rosenberg, 1968) and generation (largely by oxidative phosphorylation but also by glycolysis; Tyler et al., 1968). Although such agents as the cardiac glycosides, membrane disruptive compounds such as filipin, inhibitors of oxidative phosphorylation, and glycolysis poisons, will inhibit iodide transport, none of these is specific for thyroid tissue. Hence, these agents are not antithyroid compounds. Extensive reviews of iodide transport energetics are available (Bastomsky, 1974; Carrasco, 1993; Halmi, 1961; Wolff, 1989).
One factor whose importance has gradually emerged is the role of Na+ transport in energizing iodide transport. The first clue that Na+ flux might energize iodide transport was the demonstration that cardiac glycosides potently inhibited iodide accumulation in thyroid slices (Wolff and Maurey, 1958). Subsequently, it was shown that a sodium ion was required for iodide transport (Alexander and Wolff, 1964; Bagchi and Fawcett, 1973;Iff and Wilbrandt, 1963; Saito et al., 1982). A model was proposed in which sodium and iodide ions were cotransported in a system involving the Na+/K+ adenosine triphosphatase (ATPase) (Bagchi and Fawcett, 1973). In these systems the presence of perchlorate invariably inhibited iodide transport. With the development of thyroid cell lines, and more importantly, thyroid membrane vesicles, it became possible to analyze the details of this relation of sodium and iodide transport. O’Neill et al. (1987) andNakamura et al. (1988) showed with thyroid plasma membrane vesicles that the Na+/I−cotransport was electrogenic, with a thermodynamically downhill transport of ∼2 Na+ ions driving one iodide against its electrochemical gradient into the cell. The ouabain-sensitive Na+/K+ATPase normally mediates this gradient. These vesicle systems show a similar Km for iodide as does intact thyroid, and they are sensitive to perchlorate inhibition. As a result of such studies, the molecule responsible for the transport of iodide (and perchlorate) has been renamed the sodium/iodide symporter. Very recently the symporter molecule has been cloned and expressed into normally noniodide-transporting cells; after that, these cells now showperchlorate-sensitive iodide accumulation (Dai et al., 1996;Vilijn and Carasco, 1989); it has subsequently been cloned from human tissue (Smanik et al., 1996). There is significant similarity to other sodium-dependent transport proteins. The rat protein has 618 amino acids, the human transporter has 643 amino acids, and there is an 84% sequence identity between them. There are 12 putative transmembrane domains but neither the locus for sodium flux, the anion selectivity domain, nor the nature of their interaction has, as yet, been identified. It is to be expected that the same molecule can be identified in the other mammalian iodide concentrating tissue such as salivary gland, gastric mucosa, mammary gland, the choroid plexus, and possibly the iodide-concentrating algae such as Fucusspecies. At the same time, the symporter would be predicted to be absent or defective in patients with congenital goiter due to iodide transport deficiency (Wolff, 1983).
B. Anion Selectivity
The defining characteristic of iodide transport is its very high specificity for iodide with respect to the much more abundant chloride ion in biological systems. Nevertheless, not only iodide but several related monovalent anions are also selected. The ability to be concentrated by thyroid tissue, or to inhibit iodide transport, permits the construction of the following potency series for monovalent anion-based inhibition of iodide transport in thyroid slices (Wolff, 1964; Wolff and Maurey, 1963): TcO4− ≥ ClO4− > ReO4− > SCN− > BF4− > I−> NO3−> Br− > Cl−. Note that iodide is by no means the most highly selected anion.
Although it was long ago pointed out that this series is also the Hofmeister or lyotropic series that describes the salting-out and salting-in properties by simple ions for many proteins by concentrated solutions of neutral salts (Baumann and Metzger, 1949; Baumann et al., 1956; Collins and Washabaugh, 1985), the molecular basis of this effect is still debated. It is not clear whether this anion sequence, measured at very high concentrations, has any necessary mechanistic relation to what goes on, at low concentrations, in the thyroid. Thus, it is important to determine which solution property of the anions determined this series, e.g. crystal radius, hydrated radius, hydration enthalpy, charge density, etc. Strong base anion exchange resins (usually a large cation with weak field) exhibit a marked preference for ClO4− (compared, for example, to Cl−), thus it seemed likely that selectivity for iodide or perchlorate in the thyroid gland might be based on an ion exchange mechanism using a large cation such as a quaternary amine, e.g. arginine (Wolff, 1989). What does this relationship to anion exchange resins imply? The pioneering studies on the chemical basis of cation selectivity by Eisenman (1961, 1962) made it possible to apply equivalent arguments to anion selectivity (Wright and Diamond, 1977). A simplified view of the assumptions of this theory are: (a) Hydration energy (rather than size) is a prime determining factor for selectivity; (b) electrostatic interactions may or may not be outweighed by the hydration energy—that is, the balance between these two forces determines selectivity. The free energies required to remove water from both the fixed cation and the anion determine the total hydration energy; however, because the cation value remains constant for the leaving and entering anion, the difference between the two anions determines the selectivity. Thus, the largest anion will have the highest affinity for the exchanger because it has the lowest hydration energy. By the same token, the radius of the exchanger site will determine its field strength and, hence, the importance of coulombic interactions. The selectivity sequence depends, therefore, on the relation between changes, from one anion to the next, in the hydration energy and the number of water molecules to be desolvated and to changes in electrostatic energy. It can be shown that such manipulations (Reichenberg, 1966) readily explain the above selectivity series, and the selectivity ratio is ≫1 when comparing perchlorate to iodide and closer to one (less selective) in membrane vesicles (O’Neill et al., 1987), which may have lost some selectivity during preparation. An instructive example of the relation between anionic hydration and electrostatic forces was calculated by Reichenberg (1966)and was compared with thyroid gland selectivities by Wolff (1989). The anion selectivity of Amberlite IRA400, a strong base (weak field) anion exchanger, is explained neither by the conductances nor the coulombic interactions. On the other hand, inclusion of the hydration enthalpies to calculate the selectivities will predict the series correctly. Not all of the hydration shell need be displaced during binding to the exchanger, and the selectivity sequence is maintained when 0.6 (but not 0.5) of the hydration enthalpy remains in the calculation. The excellent correlation between the ion exchanger and the thyroid selectivity strongly suggests that such an exchange mechanism determines selectivity in thyroid tissue. Although the functional ion size is related to the above forces, one should remember that it mayper se influence permeability, as is the case in many cation channels. Further details can be found in (Wolff, 1989; Wolff and Maurey, 1962, 1963).
III. Chemistry, Metabolism, and Distribution of Perchlorate
A. Chemistry
Certain chemical properties of perchlorate are important to its usefulness in biological systems and these will be reviewed here. Salts of perchloric acid (the strongest of all common inorganic acids) have been widely used in analytical chemistry: salts of the smaller cations are very soluble, whereas salts of the larger univalent cations K+, Rb+, and Cs+ are less soluble, and perchloric acid has been used in the analysis of K+ or the removal of this cation from biological materials. Nevertheless, KClO4 is sufficiently soluble for clinical use. The anion is a tetrahedron with the four oxygen atoms at the corners and the chlorine atom at the center. The partial molal ionic volume is 44.5 ml/mol at 25°C compared with 36.7 for iodide. The Cl-O bond length is 1.44 to 1.51 Å, and the O-O distance is 2.43 Å (Kugler, 1969). The ionic radius of the iodide ion is 2.16 Å.
Perchlorate is a good oxidizing agent in reactions of the type:
In fact, however, perchlorate is stable under conditions where it is used in biological systems, i.e., dilute solution, avoidance of elevated temperatures, and neutral pH.
B. Distribution and Metabolism of Perchlorate
Extensive studies on the pharmacology of perchlorate and the other Hofmeister anions have been carried out by Eichler and Hackenthal, particularly with respect to excitable membranes (1962). Anbar et al. (1959) and Lewitus et al. (1962) used labeled perchlorate to study its possible accumulation in the thyroid. Its radioactive properties are:36Cl (production 35Cl (n, γ), half-life 3 × 105 years, β− 0.71 Mev (98%), electron capture 1.14 Mev (2%)). They found concentration of36ClO4−in rat and rabbit thyroids but, because of the low specific activity, the thyroid to medium concentration ratios of [36ClO4−] did not exceed 4 to 5. Peak uptake by the thyroid occurred at 4 to 6 hours (Goldman and Stanbury, 1973) and thyroid-stimulating hormone (TSH) increased the tissue/medium ratio of [36ClO4−].Chow et al. (1969, 1970), using smaller ClO4− concentrations, obtained much higher relative thyroid concentrations of perchlorate in rats and guinea pigs such that the thyroid/medium or serum ratios for [ClO4−] approached those for iodide. Moreover, they showed that unlabeled perchlorate or iodide had comparable inhibiting effects on the tissue/medium concentration ratios. On the basis of histological measurements and inulin and sulfate spaces, it was possible to determine the intrathyroidal distribution of36ClO4−(Chow et al., 1970). It was found that there was active transport of perchlorate from the exterior into the cell and additional further transport from the cell into the follicular lumen that may be facilitated because it exceeded the36Cl− distribution considerably. These results are comparable to the distribution of iodide within the thyroid. The ratios of the relative concentrations for36ClO4−in the different cell compartments at the lowest dose of36ClO4−that could be used were:
Perchlorate is eliminated from the body primarily in the urine. Peak blood levels after oral intake are reported to occur at 3 h. The halftimes that have been determined for the rat vary from ∼8 h (95% in 60 hours) (Eichler and Hackenthal, 1962), <8 h (Kutzim et al., 1980), to ∼20 h (Goldman and Stanbury, 1973). Despite the high redox potential (see Section III.A.), perchlorate appears to be excreted virtually unchanged both in the rat (Eichler and Hackenthal, 1962) and in man (Anbar et al., 1959). In the latter experiments, it was shown with doubly labeled perchlorate—36Cl18O4−—that the oxygen atoms of perchlorate do not undergo significant isotopic exchange in vivo, and only ∼1 to 3% of the36Cl appears as ClO3− or Cl− in the urine.
IV. Transport Effects of Perchlorate
A. Competition for Iodide
The paradigm for studying perchlorate effects derives from earlier studies using thiocyanate ion. The first hint that anions like thiocyanate or perchlorate might act competitively on iodide transport came from an early study by Astwood (1943) that showed that SCN−-induced goiter in rats was prevented by extra iodide administration. Franklin et al. (1944) found that SCN− blocked iodide accumulation in the thyroid and Vanderlaan and Vanderlaan (1947) showed that iodide ion accumulated in propylthiouracil-blocked thyroids could be readily discharged by SCN−. A more rigorous demonstration of SCN− competition with iodide was provided in mice by Wollman (1956). However, SCN− was shown to be metabolized in the thyroid and it was moderately toxic. Hence, the discovery by Wyngaarden et al. (1952, 1953), that perchlorate ion produced the same effects but was substantially more potent than SCN−, proved to be very important. These authors first showed that perchlorate markedly inhibited iodide accumulation by the rat thyroid and could discharge accumulated iodide in propylthiouracil-blocked glands. It was later shown that perchlorate effects could also occur in the unblocked thyroid (Ingbar and Freinkel, 1956). The competitive nature of the inhibition (i.e., no inhibition at extrapolated infinite iodide concentration) was established using thyroid slices and analysis by double reciprocal or Dixon plots (Wolff and Maurey, 1962, 1963). Similar results were obtained in simple systems such as isolated thyroid cells and vesicles (O’Neill et al., 1987). When the system is truly competitive, then iodide should inhibit perchlorate accumulation just as the latter inhibits iodide accumulation. Attempts to show this were made with36ClO4−and excess iodide inhibited this accumulation (Anbar et al., 1959;Lewitus et al., 1962). Because of the specific activity problem with perchlorate, attempts were made to demonstrate the reciprocal competitiveness with iodide by use of high specific activity anions, TcO4− and ReO4−, that use the same symporter. These studies showed that the Kmconstants (anion accumulation) and Kiconstants (inhibition of accumulation of one anion by another) were identical within the error of the method (Wolff and Maurey, 1962).
There is little or no effect on organification of iodide by perchlorate (e.g. Alexander, 1959, and many other in vitro studies). What little iodide enters the gland appears to be organified normally (Pitt-Rivers et al., 1958). Perchlorate thus provides a useful agent for the study of iodide transport that is not complicated by effects on later metabolic steps.
B. Locus of the Perchlorate Effect
Because of the complex anatomy of the thyroid follicle, it remains to be established where perchlorate inhibition is exerted. One of the loci is at the inward, active, movement of iodide across the basolateral membrane., i.e., the symporter (fig.1). The perchlorate effect on iodide accumulation is seen immediately upon entry of labeled iodide, i.e., at a time when not enough iodide would have accumulated to permit significant effects from alterations in exit rate. This basal location of active transport has been shown in intact thyroid tissue, thyroid cells, monolayers, and in thyroid membrane vesicles (Nilsson et al., 1990, 1994; O’Neill et al., 1987; Wolff, 1964; Wollman, 1995). On the other hand, the thyroid is, at least, a three compartment system, and much of the iodide in the intact thyroid is stored in the follicular lumen; one must, therefore, ask whether perchlorate can also act on cell to lumen transport in the thyroid. The work of Chow et al. (1969,1970) showed that, as for iodide, perchlorate accumulation occurred in two steps: an electrochemically “active” step across the basolateral membrane and an electrically downhill step across the apical membrane into the lumen; however, because the perchlorate distribution into the lumen was greater than that of chloride, more than simple diffusion appears to be involved. The fact that in polarized thyroid cell monolayers this basolateral step for iodide transport can be stimulated by TSH is in agreement with the above interpretation (Nakamura et al., 1988, 1990; Nilsson et al., 1990,1994).

Scheme of iodide transport in a thyroid follicle (modified from 184). Abbreviations are: Symp = Na/I symporter;r = receptor; Gs = activating G protein; AC = adenylyl cylase; TPO = thyroid peroxidase; EGF = epidermal growth factor. Inhibitors are italicized.Dashed lines indicate direct TSH effects on iodide movement. (*) = rapid efflux across the apical membrane; (**) = rapid efflux from lumen to the exterior.
Efflux of iodide from the lumen to the circulation requires crossing the same two membranes, as in influx, but in the reverse order. We know from early clinical experience that perchlorate promotes a very rapid and nearly complete discharge of iodide from propylthiouracil-treated glands (discussed below). Several kinetic papers have provided additional evidence for perchlorate-mediated increases in iodide efflux from the thyroid gland (Scranton and Halmi, 1965; Surks, 1967). Rocmans et al. (1977) found that all four transmembrane rates constant had to be adjusted to obtain satisfactory fits in their kinetic simulations. They proposed that perchlorate accomplished this by stimulating counter transport of iodide, but perchlorate does not induce counter transport in reconstituted thyroid membrane vesicles (Saito et al., 1982, 1989). In vitro experiments with thionamide-blocked sheep thyroid slices revealed a two phase efflux of iodide. Both components could be accelerated by perchlorate (fig. 2A). The cellular locus of these two phases is not established. The results are similar to those of sodium ion deprivation (with choline chloride) but are slower when ouabain is used to block the Na+/K+ ATPase (Salabéand Wolff, unpublished results). This sodium effect had been shown previously (Iff and Wilbrandt, 1963). This discharge should be compared with the classical study on perchlorate discharge in humans (fig. 2B), where the thionamide-blocked thyroid was loaded with radioiodide and measured externally. By current standards, very small doses (10 mg) of perchlorate achieved substantial discharge of thyroidal radioactivity, and 100 mg gave virtually complete discharge (Stanbury and Wyngaarden, 1952). This has become the standard test for organification defects as discussed in Section VIII.

Perchlorate induced discharge of thyroidal iodide from sheep thyroid slices in vitro and human thyroid gland in vivo.A. Experiment of Dr. Helga Salabé (1967).131I− -prelabeled slices blocked with propylthiouracil were placed into a large volume of medium to minimize recirculation of the released iodide. (○) = control; (•) = 1 mm ouabain; (▴) = 1 mm KClO4; (□) = 150 mm choline chloride to replace NaCl;B. External counting of a patient’s methimazole-blocked (30 mg) thyroid given a tracer dosage of 131I− at zero time. Arrows indicate oral administration of KClO4. Reproduced with permission (from Stanbury and Wyngaarden, 1952).
C. Stimulated Efflux?
Can perchlorate-induced iodide discharge from the thyroid lumen be explained by stimulation of efflux, by inhibition of influx alone, or by both? At which of the four transmembrane processes does perchlorate act? Consider the following:
The thyroid cell has a ∼50 mV negative potential with respect to either the basal or apical compartment (Woodbury and Woodbury, 1963). Thus the interstitium and lumen are roughly isopotential (fig. 1). Reduction of the potential, as is thought to result from TSH stimulation, has not been shown to follow exposure to perchlorate but will be important to measure (Williams, 1966).
Iodide movement into the cell from either direction is against an adverse electrical gradient. Across the basolateral membrane there is generally also an adverse concentration gradient; sodium flux and the Na/I symporter overcome this thermodynamic barrier and perchlorate is known to interact here. This will certainly block reentry of external iodide, but is it enough to account for perchlorate discharge of iodide?
Transport out of the cell into the lumen is electrically downhill, but high concentration gradients of lumen/cell iodide (>ten-fold) could, in principle, overcome the favorable potential according to the Nernst relation.
In the reverse direction, movement of iodide from lumen to cell is against the same potential difference, but this may, in contrast to point 3, be overcome by the high concentration gradient for iodide from lumen to cell; it may also be the rate-limiting step for perchlorate-induced iodide efflux. The gradient could be maintained by a very rapid leak of iodide from the cell to the exterior. This is, in turn, maintained by perchlorate-induced inhibition of reentry of iodide from the exterior into the cell, but it remains to be seen whether this is the only effect of perchlorate. Perchlorate added to the apical side of a polarized thyroid cell monolayer is substantially less effective than when added to the basolateral side (Nakamura et al., 1988, 1990). Moreover, perchlorate rapidly increases the secretory response of the thyroid to TSH, including both iodide and iodothyronines (Rousset et al., 1977), and TSH increases iodide efflux before it increases iodide influx (Halmi, 1961); this suggests that additional control points for perchlorate action may exist. We are left then with a readily demonstrated stimulation of iodide efflux from the thyroid produced by perchlorate, not all of which can be readily explained by competitive inhibition of the symporter for influx across the basolateral membrane (Halmi et al., 1960).
V. Anomalous Antigoitrogenic Effect of Perchlorate
In experimental animals, perchlorate leads to all the reversible effects expected from the blockade of thyroid hormone synthesis: goiter with histological signs of thyroid stimulation that fails to occur in hypophysectomized animals; thyroidectomy cells in the anterior pituitary; the corresponding changes in thyroid hormone and TSH levels; decrease in tissue QO2, etc. (Enneker et al., 1958; Krüskemper and Kleinsorg, 1954). All these effects are reversed upon cessation of perchlorate administration or with T3 or T4. Nevertheless, perchlorate goiters are not as large as those resulting from propylthiouracil treatment. In the absence of significant organic iodine formation, goiters resulting from the feeding of ClO4− are about half the size of goiters produced with propylthiouracil. Moreover, the simultaneous feeding of ClO4− together with propylthiouracil reduces the size of the goiters obtainable with propylthiouracil alone to a size characteristic of the anion (Alexander and Wolff, 1966). These findings have been confirmed in several laboratories (Jolin et al., 1968; Kahl and Bobek, 1971; Matsuzaki and Suzuki, 1981; Ortiz-Caro, et al., 1983).
Efforts to explain this anomalous effect of perchlorate have met with little success:
(a) It has been known for a long time that large amounts of iodide may reduce but not normalize thyroid size of animals fed thionamide drugs (Alexander and Wolff, 1964; Gassner et al., 1950;Goldberg and Wolff, 1954; Mackenzie, 1947; Milne and Greer, 1962). These changes may be accompanied by modest reductions in the histological signs of thyroid stimulation. It is difficult to see how perchlorate can promote a small iodide leak leading to limited hormone synthesis.
(b) Propylthiouracil inhibits the peripheral conversion of T4 to T3 but these effects are not produced by the perchlorate. Thus, when the total goitrogenic effect depends on both the intrathyroidal and extrathyroidal effects of drugs, hypothyroidism, and, hence, goitrogenesis, might be less severe with perchlorate despite the fact that the measured TSH levels are the same after both drugs (Alexander and Wolff, 1964; Jolin et al., 1968,1970; Matsuzaki and Suzuki, 1981). A variant of this explanation stems from the studies of Yamada (1967, 1968), who suggested that perchlorate interfered with the thyroxine plasma protein interaction, thus raising free thyroxine levels and diminishing the goitrogenic stimulus to the pituitary thyrotroph. A direct demonstration of T4-displacement was not provided.
(c) Jolin and coworkers (1968, 1970) found that thyroid weight of rats in response to antithyroid agents varies linearly with the circulating insulin level, and that perchlorate treatment is associated with lower insulin levels than is propylthiouracil. Exogenous insulin could restore goitrogenesis to the propylthiouracil level when ClO4− was present provided it was administered early enough during goitrogenesis and the adrenal glands are intact (Jolin et al., 1974). An additional factor is the enhancement by perchlorate of TSH sensitivity in the mouse Mackenzie assay (Rousset et al., 1977). Finally, glucose-6-phosphate dehydrogenase levels in thionamide-treated rat thyroids are reduced by perchlorate whereas ornithine decarboxylase is not (Matsuzaki and Suzuki, 1981). It may be concluded, therefore, that perchlorate possesses antigoitrogenic effects in the rat but that the mechanism of this anion effect is complex and remains to be elucidated.
VI. The Clinical Use of Perchlorate
A. Treatment of Thyrotoxicosis
Preliminary clinical studies by Godley and Stanbury (1954) on 24 patients with Graves’ disease clearly indicated that such treatment effectively controlled the disease. Although the use of perchlorate in the treatment of thyrotoxicosis was started in the Unites States, the main early experience with this compound was accumulated in Britain (Crooks and Wayne, 1960; Crooks et al., 1960; Trotter, 1962) and Germany (Krüskemper, 1960a,b, 1962; Wenzel and Lente, 1984). In Germany, iodophobia is so common among some thyroidologists that perchlorate was considered as a possible KI alternative for protection against environmental radioiodine contamination. For various reasons (potential toxicity, short duration of action, different mechanism of action, etc.) this does not seem to be justified. In addition, perchlorate is used in Germany in conjunction with thionamides for prophylaxis against iodine excess derived from coronary angiography (see Fritzsche et al., 1993 for refs.). Such precautions are not generally taken elsewhere. The role of perchlorate in the treatment of thyrotoxicosis can be viewed as having come in two waves. The first enthusiastic period lasted 6 to 7 years and saw well over 1000 (published) patients treated successfully with a side reaction rate not very different from what was seen with thionamides. The appearance between 1961 and 1966 of seven cases of aplastic anemia put a virtual stop to the use of perchlorate, except for single dose-use in perchlorate discharge tests and as an adjunct to pertechnetate scanning. However, with the massive use of amiodarone, a second wave of perchlorate use has been initiated with the lessons learned from earlier reactions kept well in mind.
Initially 400 mg/day taken orally were used in divided (4 to 5/day) doses. The use of frequent daily doses seemed to be important because the blocking effect on iodide uptake was short lived. This led to good, although not very rapid, control of thyrotoxicity with no major toxic reactions save for gastrointestinal disturbances alleviated by dosing with meals (Godley and Stanbury, 1954). It was subsequently found that this dose did not control thyrotoxicity in some patients, and larger doses were initiated (Crooks and Wayne, 1960; Crooks et al., 1960;Trotter 1962). With 600 to 1000 mg KClO4/day in five divided doses, control was good, generally occurring in 3 to 8 weeks but was attained somewhat more slowly than with methylthiouracil. In some clinics, KClO4 treatment was started with large initial doses followed by reduction to maintenance levels of 400 to 600 mg/day or to even lower levels (Krüskemper, 1960a,b;Wenzel and Lente, 1984). In a different study using even larger daily doses (1.5 to 2.0 g/day) response was faster (9.4 weeks to remission) than with carbimazole (12.3 weeks), but the number of side reactions was decidedly greater than observed on a regimen of 600 to 1000 mg/day.
One critical difference between thionamides and perchlorate is the danger of using iodides in conjunction with perchlorate, e.g. for the reduction of thyroid vascularity preoperatively. Although this is safe for the thionamides, the fact that iodide and perchlorate are competitive for transport mitigate against its use for the reduction of vascularity much favored by surgeons. The extreme vascularity in patients prepared with perchlorate caused technical difficulties at surgery, hence thionamides, plus iodide is a safer method of preparation for surgery (Cook and Hawe, 1960; Godley and Stanbury, 1954). For the same reason, other iodine-containing medications should be avoided, when possible. Despite the many chemical similarities between the two anions, this is a clear difference from iodide and suggests that the iodide effect on thyroid vascularity may involve the formation of an organic iodine inhibitor not possible with perchlorate (Wolff, 1996).
Perchlorate has been used also when sensitivity to one of the thionamides developed. Combined therapy with perchlorate and thionamide, with the aim of permitting the use of lower concentrations of perchlorate, has been successful (e.g. Blørstad and Vogt, 1962;Røkke and Vogt, 1968); the time to euthyroidism was not appreciably shortened, side reactions are more difficult to assign to a specific drug, and the use of an alternative therapy is lost. Nevertheless, combined therapy has been successful in the occasional cases of iodide-induced thyrotoxicosis after angiography (Fritzsche et al., 1993), in functioning metastatic thyroid carcinoma (Joseph et al., 1994), and particularly in amiodarone-induced thyroid dysfunction (see Section VII.).
B. Perchlorate Toxicity
Experimental studies on perchlorate side reactions in rats at relative dosages much larger than those used in man had shown very little evidence of toxicity (Krüskemper, 1960b), and the clinical findings during the first 6 years revealed primarily minor side effects consisting of nausea or gastritis relieved by administration of the drug with meals, rashes, occasional thyroid enlargement, and leukopenia that was difficult to differentiate from that occurring naturally with thyrotoxicosis. In a review of 881 cases, Krüskemper (1960b,1962) found these reactions to amount to 3 to 4%, less than seen with the thionamide drugs (see also Røkke and Vogt, 1968). In a more extensive review of the relative toxicity of antithyroid drugs, Trotter (1962) has classified side reactions into three groups: (a) Minor—having an overall incidence of ∼10% including drug rashes, drug fever, lymphadenopathy, or nausea. (b) Agranulocytosis with an incidence of ∼1%; several cases of the nephrotic syndrome were reported at the initial trials but not since. (c) Aplastic anemia with a low incidence. For Trotter (1962), the total incidence of reactions was 2 to 3%, substantially lower than simultaneously estimated reactions to thionamide drugs. As familiarity with perchlorate use increased, dosages were increased in an effort to speed reduction of thyrotoxicity, and it is with these doses (1200 to 2000 mg/day) that there was a large increase in side reaction, going from 2 to 3% to 16 to 18%. Agranulocytosis appeared to be less frequent (0.3% of 1200 cases including single case reports) than in 10,131 cases treated with thionamides (0.94%). The appearance of seven cases of fatal aplastic anemia in 1961 to 1966—four in Britain in 1961 (viz. Trotter, 1962), one in the United States (Krevans et al., 1962), one in Norway (Gjemdal, 1963), and one in Israel (Barzilai and Scheinfeld, 1966)—virtually stopped the use of perchlorate except for single doses used in the perchlorate discharge test (see Section VIII.). It has been popularly assumed that these cases resulted from the use of large dosages of perchlorate, but this is not the case, as all cases followed doses ≤ 1 g/day. On the other hand, most of these patients were treated for periods of 2 to 6 months before aplastic anemia was detected and duration may be a contributing factor. The fact that four of these cases occurred in a cluster raises the question as to whether the perchlorate used at that time may have been contaminated. This question was not followed up at the time. No serious side reactions have been recorded with the resurgent use of perchlorate (Bartalena et al., 1996), and one case with a 22-year maintenance dosage of 200 mg/day had no complications (Connel, 1981). Personal inquiries by Trotter in the 1960s and recently by this author (Germany, Switzerland, Holland and Italy) regarding toxicity after its use for amiodarone-induced thyrotoxicosis at lower dosages and for shorter times, (see Section VII.D.) have not brought to light any additional cases of aplastic anemia or other serious side reactions.
Finally, it must be pointed out that perchlorate readily crosses the placenta (of the guinea pig) and may produce goiter in the fetus, whose thyroids appear to be more sensitive to this anion than that of the mother (Postel, 1957). Crooks et al. (1960) have treated 12 pregnant thyrotoxic women with satisfactory results; 1 neonate had a small, transient goiter.
VII. Amiodarone
The second phase of perchlorate therapy, still ongoing, is based on the special advantages of perchlorate in cases of thyrotoxicosis caused by large iodide loads. Most of these result from the use of the antiarrhythmic agent, amiodarone, but the following arguments are equally applicable to other organic iodine compounds depending only on their half-lives and rates of deiodination. Amiodarone (2-butyl-3-[3,5-diiodo-4-(β-diethylaminoethoxy)-benzoyl]benzofuran) (fig. 3) is an iodinated benzofuran derivative containing 37.3% iodine and varying amounts of free iodide.
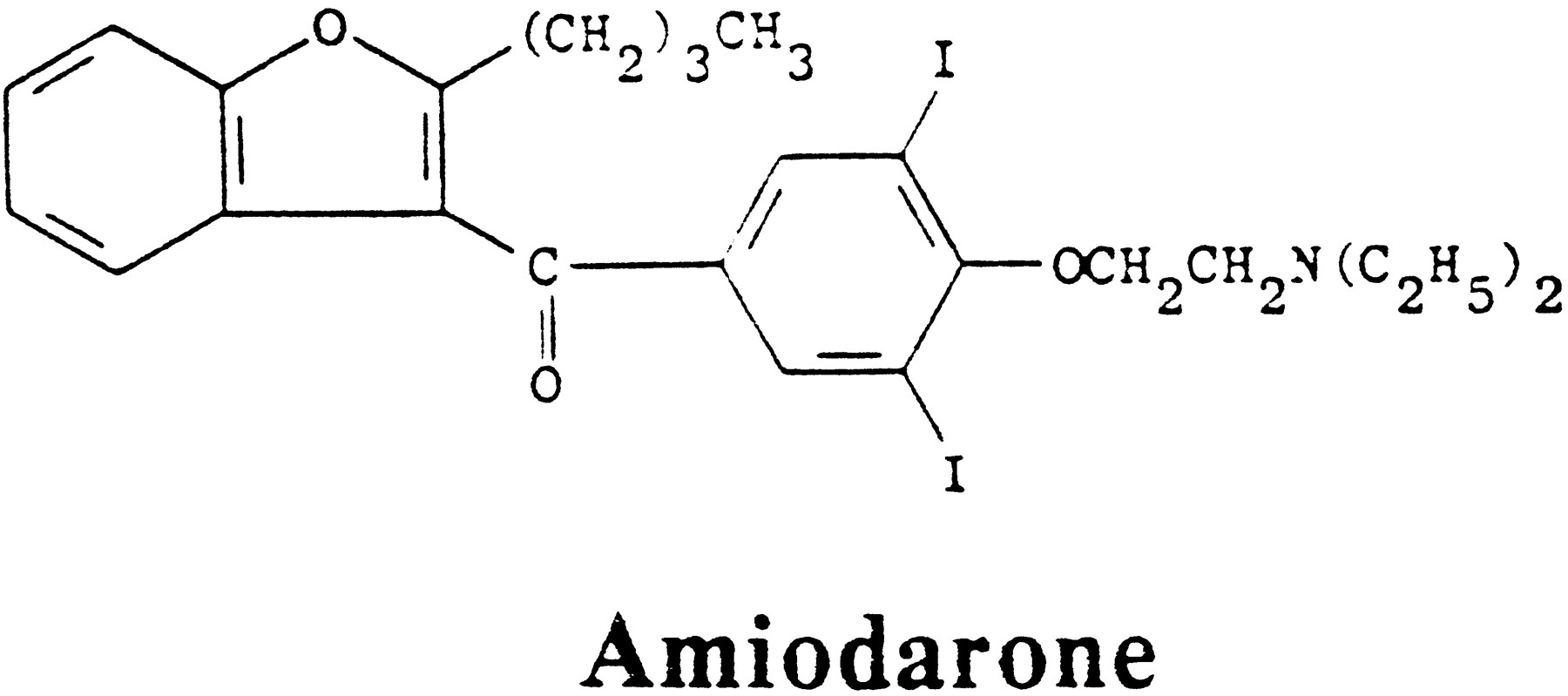
The structure of amiodarone.
The drug is stored extensively in many tissues, particularly adipose tissue, and the distribution volume averages 5000 l, leading to a biological half-life of many weeks to 8 months (Holt et al., 1983;Latini et al., 1984, Léger et al., 1984). Its blood level is chronically maintained at 0.7 to 0.9 mg/l (Rao et al., 1986), but values up to 4 mg/l have been reported (Shukla et al., 1991). In one human thyroid, the amiodarone concentrations were about 20 times higher than in blood (Smyrk et al., 1987). In other reports thyroid concentrations varied from 5 to 25 mg/kg, and 75 mg/kg for desethylamiodarone, a metabolite of amiodarone (Haffajee et al., 1985;Holt et al., 1983; Latini et al., 1984). The vast clinical literature, much of it from Europe, where the drug had been more popular than in the United States, reveals that its use is accompanied by numerous side reactions in various organ systems (Mason, 1987). Nevertheless, because the drug is believed to be the only one available for many refractory arrhythmias, these side reactions are often accepted. Among these untoward effects are three of considerable significance for the thyroid. They are: (a) a change in the peripheral metabolism of thyroxine; (b) a direct toxic effect on the thyroid parenchyma; (c) several effects resulting from the excess iodide produced by deiodination of amiodarone. It is important to remember that side effects not easily ascribed to excess iodide alone complicate diagnosis and therapy. We shall summarize these briefly before dealing with the perchlorate-sensitive effects of amiodarone-derived excess iodide.
A. 5′-Deiodinase Effects
A confounding factor in the assessment of the thyroid status in the presence of amiodarone is the fact that amiodarone interferes with the deiodination of T4 to T3 by the Type I, propyl-thiouracil-sensitive, 5′-deiodinase (Burger et al., 1976; Franklyn et al., 1985; Melmed et al., 1981; Nadamanee et al., 1989; Singh and Nadamanee, 1983), which accounts for the bulk of peripheral T3production. This results in an increase in total and free T4, an increase in rT3, and a corresponding decrease in total and free T3 in the plasma and most tissues. The Type II, propylthiouracil-resistant 5′-deiodinase of pituitary tissue and brain is apparently little affected by amiodarone. This is probably the most common thyroid abnormality present in amiodarone-treated patients but is generally not accompanied by clinical dysfunction (Newnham et al., 1988). Direct inhibition of the 5′-deiodinase by amiodarone has also been demonstrated in hepatocytes with an IC50 of ∼1 μm (Aanderud et al., 1984). Such an effect is not produced by an equivalent amount of iodide. Another confounding factor is an increase in the TSH response to thyrotropin-releasing hormone, and a second hormone-binding-related action of amiodarone or its primary metabolic product, desethylamiodarone, is the noncompetitive inhibition of thyroid hormone binding to the β1receptor and competitive inhibition to the α1receptor, which may cause local hypothyroidism (Bakker et al., 1994;Franklyn et al., 1985; Van Beeren et al., 1995). It has been suggested that some of the salutary effects of amiodarone, particularly in the heart, might be explained by this mechanism.
B. Direct Thyroid Amiodarone Toxicity
A direct toxic effect from intact amiodarone (or its monodeethylated product) has been proposed on the basis of a comparison of postmortem examination of thyroid glands of amiodarone-treated patients who developed thyrotoxicosis; they were found to have thyroid lesions whereas patients without thyrotoxicosis did not (Smyrk et al., 1987). Thyroid damage is seen in needle biopsy material (Capiello et al., 1995) and tissue culture experiments (Brennan et al., 1995). Cell damage suggests a destructive thyroiditis, as shown by decreased chromium retention, loss of tissue contents including thyroglobulin and, hence, thyroid hormones, and decrease in cyclic adenosine 3′,5′- monophosphate (cAMP) production (Beddows et al., 1989; Massart et al., 1989). The mechanism of the toxicity is not clearly understood. The lesions produced resemble the damage that is sometimes seen after exposure to excess iodide (Belshaw and Becker, 1973; Many et al., 1992). Reports that both methimazole (Chiovato et al., 1994) and perchlorate (Brennan et al., 1995) prevent thyrocyte damage produced by high concentrations of amiodarone suggest that here too excess iodide may have contributed to the lesions. However, because similar types of damage are also seen in tissues that do not concentrate iodide (Beddows et al., 1989; Chiovato et al., 1994), there can be little doubt that a direct toxic effect is one factor that has to be kept in mind when evaluating amiodarone-induced thyroid dysfunction. Such effects would obviously not be subject to perchlorate competition. As might be expected from the documented tissue damage, several examples of subacute thyroiditis accompanied by thyrotoxicosis, frequently painful, have been encountered (Roti et al., 1992, and references therein). Thyroglobulin levels may be transiently elevated but antithyroid antibodies generally are not. Some of these patients proceed to subclinical hypothyroidism and will show a positive perchlorate or iodide-perchlorate discharge test (see below). Similarly, direct toxic effects may explain the clinical course of patients not responding to perchlorate therapy for amiodarone-induced thyrotoxicosis as well as their response to corticosteroids (Bartalena et al., 1996; Capiello et al., 1995).
C. Amiodarone and Iodide Excess
Amiodarone is metabolized both by the loss of one of the ethyl groups and by deiodination, which is the chief degradation product of interest here. Approximately 10% of the organic iodine is said to be released as iodide per day (Broekhuysen et al., 1969), but it is not known in which organ the bulk of the deiodination occurs. Plasma inorganic iodide concentration was found to rise from 0.05 μm to 2.2 μm, and urinary iodide excretion increased from 0.25 μmol/mmol creatinine to 7 μmol/mmol creatinine or more (Rao et al., 1986). As a result, the absolute thyroid iodine uptake increased to a maximum of 55 nmol/hour. This, in turn, increases the thyroid iodine content as measured by X-ray fluorescence and tends to be greater in patients who become hyperthyroid than in those who become hypothyroid (Fragu et al., 1988; Jonckheer and Deconinck, 1982;Léger et al., 1983, 1984). However, the majority of amiodarone-treated patients have functioning adaptive mechanisms that block or reduce the massive iodide uptakes (Fradkin and Wolff, 1983), and generally only those patients that develop or have developed thyroid dysfunction show these increases. Two forms of dysfunction are observed, roughly as a function of the iodide intake of the population. With adequate dietary iodine supplies, the more common problem is hypothyroidism, whereas in iodine-deficient regions, iodide induced thyrotoxicosis is more common. Put another way, the ratio of hyper/hypothyroidism is <1 in iodine-sufficient areas and >1 in iodine-deficient areas. The incidences reported for thyroid dysfunction vary widely, often because the series are small, from 2 to >30% of amiodarone-treated patients (Harjai and Licato, 1997; Martino et al., 1984; Nadamanee et al., 1989; Trip et al., 1991). An unknown fraction of these patients may have become so because of the direct effects of the drug on the thyroid rather than by iodine excess. Two types of amiodarone effect are important to distinguish (see below) because therapy in cases with damaged thyroid parenchyma will be different. Nevertheless, the success of perchlorate therapy in many of these cases (see below) strongly implicates excess iodide as the cause of the hyperthyroidism.
D. Amiodarone-Induced Thyrotoxicosis
As might be expected, many of the effects of amiodarone resemble those induced by other sources of iodide. Hundreds of cases have been recorded, particularly in Europe and particularly from Martino and coworkers (Fradkin and Wolff, 1983; Martino et al., 1984, 1986a,b,1987a,b, 1988). Amiodarone-induced thyrotoxicosis (AIT) may appear, often quite suddenly, early or after many years of therapy with amiodarone (Newnham et al., 1988) and may appear many months after withdrawal of amiodarone therapy. It occurs in patients with or without known preexisting thyroid disease. The sex ratio is 3 to 1 males>females; this is in excess of the 2 to 1 ratio of amiodarone-treated patients who remain euthyroid. AIT is not generally accompanied by changes in antibody levels unless preexisting thyroid disease was present (see also Section VII.B.). Because of the large, long half-life, stores of amiodarone, AIT is more difficult to treat than other forms of iodide-induced thyrotoxicosis. Drug withdrawal occasionally is sufficient but generally remission is slow (Martino et al., 1987a), and some cardiac situations do not permit drug withdrawal. The importance for avoiding thyrotoxicosis in patients with cardiac arrhythmias is self evident, and fatalities from uncontrollable thyrotoxicity have been recorded despite discontinuance of the drug. Occasionally, AIT subsides despite continued therapy. For patients in whom amiodarone cannot be discontinued, examples exist of successful therapy of AIT during continued amiodarone therapy (Reichert and de Rooy, 1989; Trip et al., 1994; Wémeau et al., 1991). The traditional treatment with thionamide drugs such as propylthiouracil or methimazole has had rather limited success. This is perhaps understandable when the following is considered: Assume that thionamide drugs inhibit hormone formation by 95% under normal iodide loads. If now the iodide load is increased by say thirty-fold (which seems quite probable from what has been measured; Rao et al., 1986), then more organic iodine and thyroid hormone will be synthesized and stored despite what would appear to be adequate therapy for routine Graves’ disease. It is for this reason that attempts were made to reduce the excess thyroidal iodide by preventing entry via the symporter and by discharging iodide that might accumulate in the gland from intrathyroidal deiodination.
Martino and coworkers (1986a) compared three treatment modalities in patients with AIT in whom the amiodarone had been discontinued: no treatment, methimazole (40 mg/day) alone, and methimazole plus KClO4 1g/day. The unequivocal conclusion was that only the combined drug treatment controlled thyrotoxicity in allcases, and that the time required for attainment of the euthyroid state was greatly reduced (to 1 month or less) compared to those cases that did respond to other treatments. After 40 days, perchlorate was stopped and control was possible with methimazole alone or without further treatment. With combined treatment the return to euthyroidism was a function of the state of the gland before amiodarone: with dysfunction it took 15 to 90 days, without it, it took 6 to 55 days (Martino et al., 1987a). The difficulty in treating thyrotoxicosis in the presence of an iodide load has been noted previously, as has the success of treating Graves’ disease rapidly with combined thionamide and perchlorate (Léger et al., 1984; Røkke and Vogt, 1986). It is of interest that the urinary iodine output in the patients in whom amiodarone had been withdrawn showed a brisk increase in the first week or two after institution of the combined therapy. Perchlorate in the absence of amiodarone will also do this (De Groot and Bühler, 1971). These results have been confirmed (e.g. Newnham et al., 1988). In some series the response to combined therapy is slower than above, and there are occasional patients that are refractory, possibly due to direct toxic amiodarone effects on the thyroid parenchyma (see above). Perchlorate dosages have varied between 600 and 1000 mg/day but not more because of the fear of serious side reactions. We are not aware that a minimum effective dosage has been established when used in combination with thionamides.
An unknown fraction of these patients may have become thyrotoxic because of the direct effects of the drug on the thyroid rather than by iodine excess. The latter may not respond to perchlorate and are treated with corticosteroid as for subacute thyroiditis. The distinction between these two types of AIT is now being made and has been classified in table 1 (Bartalena et al., 1996).
Comparison of type I and type II amiodarone-induced thyrotoxicosis
E. Amiodarone and Hypothyroidism
Inhibition of thyroid function by excess iodide leading to hypothyroidism and/or iodide goiter has been studied extensively in the past (reviewed in Wolff, 1969). It appeared to be more common in areas of sufficient dietary iodine intake and so it is with amiodarone (Harjai and Licato, 1997; Hawthorne et al., 1985; Martino et al., 1987a,b). Because hypothyroidism is more common in patients with preexisting thyroid dysfunction, many thyroidologists assume that iodide-induced hypothyroidism is presumptive evidence for the unmasking of a preexisting thyroid disorder such as preclinical Hashimoto’s thyroiditis. Although this may be the case after amiodarone, hypothyroidism can occur in apparently normal thyroid glands as best as can be established. Of course, when amiodarone induces destructive thyroiditis, it is likely that hypothyroidism may eventually occur (see above). Hypothyroidism after amiodarone can be so severe as to lead to myxedema coma (Mazonson et al., 1984), or it may be merely a laboratory diagnosis (Roti et al., 1992, 1994). An interesting finding is that there may be uninhibited or even increased radioiodine uptake (Martino et al., 1988; Wiersinga et al., 1986). Such patients frequently show a positive perchlorate discharge test indicating incomplete organification of iodide.
The obvious therapy is exogenous thyroid hormone and this has been widely used. Care has to be taken that the amiodarone-induced inhibition of the conversion of T4 to T3 is taken into account. It is, of course, especially important to avoid overdosing with T4or T3 in patients who already have arrhythmias. Of greater interest for the present review is the counter-intuitive finding that perchlorate can discharge enough iodide and/or block enough iodide entry into the gland to restore clinical and biochemical euthyroidism in many patients in whom hypothyroidism was induced by amiodarone. This type of effect was first noted many years ago by Raben (1949) who observed that the inhibition of organic iodine formation caused by excess iodide could be reversed by thiocyanate. In six patients with amiodarone-induced hypothyroidism, 1 g of potassium perchlorate for 9 to 34 days restored euthyroidism with decreased TSH levels and increased FT4I (Martino et al., 1986b), and was maintained for some time. In others it reverted in 5, 47, and 73 days after cessation of perchlorate therapy but could again be normalized with repeat perchlorate therapy. Similar results have been obtained by others (Van Dam et al., 1993; Wémeau et al., 1991).
How can this paradoxical effect of perchlorate be explained? Excess iodide inhibits many parameters of thyroid function from iodide transport, to cAMP generation, to hormone synthesis, etc. (reviewed inWolff et al., 1989). In all cases, these effects occur only when oxidation of the accumulated thyroid iodide is permitted. This has led to the postulate that an iodinated organic inhibitor is formed. Recently, several iodinated lipids, iodolactones derived from arachidonic acid, and 2-iodohexadecanal have been identified in the laboratory of Boeynaems in Brussels (viz. Wolff, 1996) as potential inhibitor candidates.
One obvious problem with this proposal would be that the reduction of iodinating species might decrease both protein and lipid iodination equally. The important discovery that lipid iodination required ∼10 times more iodide than protein iodination (Pereira et al., 1990) offers a possible explanation for the salutary effect of perchlorate in amiodarone induced hypothyroidism: perchlorate will disproportionately affect lipid iodination while still permitting protein iodination. This is still hypothetical but offers a pathway for thinking about the apparently paradoxical effect of perchlorate.
VIII. The Perchlorate Discharge Test
A. Kinetic Considerations
The significance of the perchlorate discharge test is best understood by considering the following simplified scheme for the early steps of the iodine pathway in the thyroid. In an unblocked gland, the simplest mode for depicting organic iodine formation is:
1. k1 Effects.
When the thyroid is stimulated but k1 is increased more than k2 or k3, as may happen in Graves’ disease (see below), then dischargeable intrathyroidal iodide may accumulate. Reduction of thyroid mass by surgical or radioiodine ablation may also lead to a buildup of the iodide pool. This may be the result of TSH stimulation of the remaining tissue and the consequent increase in iodide transport as was shown long ago for induction of iodide inhibition by low iodine diets in the rat thyroid (Wolff, 1969). Because the total entry of iodide into the thyroid is a function of [I−]k1, increased iodide loads [I−] i.e., increased serum iodide levels, may increase iodide accumulation in the thyroid with or without increases in k1. This will eventually be in excess of what can be handled by the subsequent iodination steps. This occurs with iodine-rich drugs or diet (seaweed) and does not necessarily imply an iodide effect on the oxidizing system. The relative iodide load effect is also the basis of the iodide-perchlorate test that has greater sensitivity than perchlorate alone for the detection of organification defects (see Section VIII.B.).
2. k-1 Effects.
We are not aware of any examples of reduced iodide efflux leading to iodide accumulation in the gland, but such an effect cannot, in principle, be ruled out even though the main effect of perchlorate is on iodide influx (decreasing k1).
3. k2 Effects.
Oxidation of iodide to reactive forms such as I° or I+ requires, as a minimum, thyroid peroxidase (TPO) and a peroxide-generating system. Absent or defective peroxidase (mutations, mislocation, absence of protoporphyrin IX, etc) must lead to the accumulation of dischargeable iodide in the gland as seen, for example, in certain congenital goiters. The classic example of such a block is the attack on the oxidizing system by thionamide drugs. This is, in fact, the finding that started the discharge tests, first with thiocyanate (Stanley, 1949; Stanley and Astwood, 1948) and then with perchlorate (Stanbury and Wyngaarden, 1952). Absence of peroxide could also lead to iodide accumulation. We are aware of only one example of defective peroxide-generating systems (Kasukabe, 1975).
4. k-2 Effects.
In theory, reduction of the oxidized iodine species to iodide, by excess intrinsic thiol groups in the thyroid, for example, might contribute to an enlarged iodide pool. This would resemble the end result of thionamide exposure. We know of no data that has actually demonstrated this. In this regard it is of note that excess iodide leads to an increase in thyroid glutathione (Allen, 1993).
5. k3 Effects.
Oxidized iodine requires a protein acceptor (usually thyroglobulin). Some examples of defects in thyroglobulin have been reported and blamed for positive discharge tests, although the reason for the inability to iodinate at least some of the remaining nonhormonogenic tyrosyl residues has remained obscure. Decreased substrate (thyroglobulin) concentrations also seem unlikely because tyrosyl residues of other proteins in the thyroid could serve as alternate acceptors.
The perchlorate discharge test has been carried out in many variations. Tracer 131I or 123I is given 1 to 2 hours before perchlorate, neck counts are taken (preferably corrected for neck background), and then 200 to 1000 mg of KClO4 are given orally. It is not clear that the larger dosages are necessary. At early intervals thereafter (15 to 60 minutes, occasionally 120), neck counts are repeated. Timing is important and should be short to minimize counts from organic iodine in partial defects. Discharge of isotope from the neck is usually rapid in positive cases and may or may not be followed as a function of time to see how low the residual neck count will get. When discharge is slow, e.g., significant only after >2 hours, consideration of additional factors should be entertained. The criteria for positivity vary widely in different clinics and range from a loss of >10% to >30% of the radioactivity accumulated at the time of perchlorate administration. This is, in part, a function of the precise timing and the absolute radioiodine uptake (dietary iodine) of the region (Martino et al., 1984; Trip et al., 1991). The intravenous perchlorate test can be carried out at much shorter time intervals, e.g., 10 minutes after radioiodine and is felt to be significant with 1 to 2% discharge (Gray et al., 1973; Hilditch et al., 1982). It is uncertain whether much is gained by this variation.
B. The Iodide-Perchlorate Test
As detailed in the previous section, administration of an iodide load can increase the intrathyroid iodide pool such that the capacity of the iodinating system may be exceeded more easily. This would be expected to expose marginally sufficient oxidizing systems and lead to a larger fraction of positive perchlorate discharge tests but may become meaningless when the iodide load is so large as to exceed the normal iodinating capacity or provoke its own antithyroid effects. Thus, normal patients may show positive tests when the loading iodide dose reaches ≥ 1 mg (Childs et al., 1950; Stewart and Murray, 1967). In this test, iodide is given along with the radioactive tracer in doses varying from 200 to 1000 μg. The remainder of the test is carried out as above. The large variation in the incidence of positive discharge tests in Hashimoto’s thyroiditis (Morgans and Trotter, 1957;Murray and McGirr, 1960; Volpé et al., 1965) led Takeuchi et al. (1970) to compare the standard discharge test with one in which 500 μg of I− had been given with the131I. Under these conditions normal subjects still showed no discharge, whereas in Hashimoto’s thyroiditis (8 patients) the number of positive tests increased from 2 to 8. Other thyroid disorders show similarly increased discrimination for marginal defects (Friis, 1987).
C. Examples of Iodide Excess
Iodide given in excess, either as inorganic iodide or derived from organic precursors (contrast media, amiodarone, etc.) given over prolonged periods of time, leads to iodide goiter, among other problems (Wolff, 1969). Many of these patients exhibit positive perchlorate discharge tests. Although occasional thyroid damage has been demonstrated after iodide, the fact that most of these cases are fully reversible suggests that this must be a minor complication in the majority of such patients. It is not always clear when this is due simply to a load effect, [I−]k1, or indicates inhibition or various thyroid functions (Wolff, 1989). Recovery after cessation of iodide excess suggests a load effect. For amiodarone treatment, simple reflection suggests that hypothyroidism would be more likely to yield positive discharge tests than hyperthyroidism. This has proved to be the case (Martino et al., 1988; Roti et al., 1994).
Of considerable interest is the finding that patients with Graves’ disease may have dischargeable iodide pools in their thyroids (Roti et al., 1994). Although most metabolic processes are likely to be accelerated in such glands, this does not occur to the same extent in all of them and the positive perchlorate test suggests, but does not prove, that there is a proportionately greater increase in iodide transport than in organification. The possibility that this may be due to tissue damage (as with coexisting thyroiditis) should be kept in mind.
D. Examples of Organification Defects
Historically, discharge of accumulated intrathyroidal iodide was first demonstrated in thionamide-blocked glands by Stanley and Astwood (1948), who used thiocyanate to compete for iodide. When the advantages of perchlorate became apparent, this anion was substituted and proved extremely effective (as shown in fig. 2A). This figure shows nearly total discharge of label after administration of 100 mg KClO4, and substantial discharge by as little as 10 mg, leading to the conclusion that the thionamide block was virtually complete. This has been confirmed many times since. In some cases of Graves’ disease treated for prolonged periods, a positive iodide-perchlorate discharge test persists 40 days after drug withdrawal (Roti et al., 1994). It is debated whether this is a drug-induced long-term effect or persistence of the hyperthyroid state. Patients treated with lithium salts for bipolar disease often exhibit antithyroid effects and may show a positive iodide-perchlorate discharge test (Andersen, 1973). The locus of lithium action in the thyroid is not established. See table 2 for these and other abnormal discharge tests.
Positive perchlorate discharge tests
Failure of the oxidizing system of the thyroid may result from a variety of causes (Furth et al., 1967; reviewed in Lever et al., 1983;Medeiros-Neto and Stanbury, 1994). Functional thyroid peroxidase may be deficient or completely lacking as seen in congenital goiters or defective because of its inability to bind one of its substrates. There may be a defect in protoporphyrin IX binding; in certain porphyrinopathies from lead poisoning or hepatic porphyria (Haust and Meagy, 1980), there may be positive discharge tests, but these tend to show a slow release of radioiodine, and other factors may also play a role (Harjai and Licato, 1997). Thyroid peroxidase is normally bound to the apical cell membrane, and mislocation has been proposed as a possible cause of defective organification. In general, however, the structural defects in the peroxidase remain to be elucidated. Finally, a lack of thyroglobulin may be accompanied by normal or high levels of peroxidase and thereby lead to iodide accumulation. Abnormal thyroglobulin has also been implicated, but, as mentioned above, it is difficult to comprehend how all of the solvent-exposed tyrosyl residues of thyroglobulin could have become unavailable for iodination (Floyd et al., 1960). It is of considerable interest that neonatal hypothyroidism associated with a positive discharge test may be transient (Nose et al., 1985). This was ascribed to immaturity of the organification mechanism.
Pendred’s syndrome, congenital nerve deafness with goiter and a positive perchlorate discharge test (Fraser et al., 1960), is an example of a partial organification defect in which patients are usually euthyroid. One hundred years after its first description and after a study of nearly 1000 cases, the basis for the goiter and positive discharge test remains obscure. Despite a few reports of defective thyroid peroxidase, most investigators feel that the defect resides elsewhere (Medeiros-Neto and Stanbury, 1994). It has also been suggested that abnormal thyroglobulin may be associated with the syndrome, but again this is far from established. It is of interest that the presence of a positive perchlorate discharge test has become asine qua non for the diagnosis.
The most common example of positive perchlorate or iodide-perchlorate discharge tests is found in Hashimoto’s thyroiditis (Morgans and Trotter, 1957; Murray and McGirr, 1960; Volpé et al., 1965). Dischargeable iodide is sometimes absent in this disorder but more often is associated with a partial organification defect. The literature on this defect in Hashimoto’s thyroiditis is extensive and will not be reviewed here except to note that in mild cases it may require the iodide-perchlorate test to evoke a positive result. The great sensitivity of Hashimoto’s thyroiditis toward iodide in inducing thyroid blockade probably describes the same phenomenon (viz. Wolff, 1969). Despite the fact that a positive perchlorate discharge test is often observed in Hashimoto’s thyroiditis, the details of the defect are not understood. Because of the frequent presence of anti-TPO antibodies, some have assumed that the antibody may have damaged or inhibited the peroxidase in the residual functioning thyroid tissues. This has not, to our knowledge, been demonstrated. One could postulate that with slow cell damage, functions at the basolateral membrane might survive longer than the organification system, for example. Such dissociation has, in fact, been observed after irradiation of thyroid tissue (Barzelatto et al., 1962; Maloof et al., 1952), but information for thyroiditis is not available. It has also been suggested that Hashimoto thyroid tissue may iodinate thyroglobulin but not catalyze the coupling reaction to form T3 and T4 (Medeiros-Neto and Stanbury, 1994). This is, however, not readily compatible with the positive discharge tests. Women with previous postpartum thyroiditis may show persistent positive iodide-perchlorate discharge tests when they have gone through both the hyper and the hypothyroid phases, but not when they suffered only from the transient hyperthyroid phase (Creagh et al., 1994).
E. Ectopic Thyroid Tissue
Lingual thyroids may exhibit any level of thyroid function from normal to no function at all, and they may be malignant. It is thus not surprising that organification defects have been found. In 9 of 20 cases of lingual thyroid, positive perchlorate discharge tests were measured (Ferrini and Biassoni, 1966). Interestingly, in one of these with both in situ and ectopic tissue, only the lingual tissue showed a positive test. In 13 lingual or sublingual locations, the mean perchlorate discharge of 123I was 43% (Hilditch and Jackson, 1985). Pertechnetate can be similarly discharged by perchlorate from lingual thyroids or ectopic tissue, as visualized by scintigraphy (Al-Jurayyan and El-Desouki, 1997; El-Desouki et al., 1995). As with some congenital thyroids, the defect may be transient. Finally, a positive discharge test was shown scintigraphically in a follicular and spindle cell carcinoma metastasis to the skull (Valenta, 1966) (see table 2; note that some items are classified under several headings; in addition, a great many references to discharge tests are left out for reasons of space).
Finally, a less well-known use for perchlorate, not directly linked to thyroid function but illustrating the use of lessons learned from iodide-concentrating tissues, generally, such as salivary glands, choroid plexus, gastric mucosa, etc., is its use in pertechnetate (99mTcO4−) scintigraphy, for example, in the brain. Contaminating pertechnetate in organic technetium reagents tends to enter the parotid gland. Prevention of this by perchlorate improves the images (Ballinger et al., 1990). Because pertechnetate and perchlorate are both competitors for the Na/I− symporter in all tissues where it is found, perchlorate may be used to identify certain choroid plexus papillomas. The pertechnetate that has been accumulated in these structures can be discharged by perchlorate, whereas in other tumors or in the rest of the brain, labeled technetium cannot be discharged (Cleto et al., 1992). Perchlorate is similarly useful in pertechnetate investigation of gastrointestinal bleeding (Hilditch et al., 1985).
IX. Summary
Perchlorate competitively blocks iodide from entering the thyroid by an effect on the Na+/I−symporter thus preventing the further synthesis of thyroid hormone but has no effect on the iodination process itself. It is concentrated by thyroid tissue in a manner similar to iodide but is not significantly metabolized in the gland or peripherally. What is not settled is whether there are additional perchlorate effects on iodide transport. Perchlorate has a fast turnover in the body and requires frequent daily doses for therapy of thyrotoxicosis. Perchlorate appears to be substantially more effective against large iodide loads than the thionamides, and, with long-term iodide contamination, combined therapy of perchlorate (with ≤ 1 g/day) and thionamides is recommended for the more severe cases of thyrotoxicosis that may result from excess iodide or iodide-generating organic compounds, as for example with amiodarone. After ∼30 days, the perchlorate dosage can be tapered or stopped, continuing with thionamides alone. This markedly increases its safe use. Despite serious side effects during its early use, lower dosages and shorter treatment periods appear to have prevented such reactions in its recent reintroduction, mostly for amiodarone-induced thyroid dysfunction. Perchlorate can also protect against inhibition of thyroid function and the resulting hypothyroidism caused by excess iodide, presumably by reducing the formation of an iodinated inhibitor. The reduction of the iodide pool by perchlorate thus has dual effects—reduction of excess hormone synthesis and hyperthyroidism, on the one hand, and reduction of thyroid inhibitor synthesis and hypothyroidism on the other. Perchlorate remains very useful also as a single dose application in tests measuring the discharge of radioiodide accumulated in the thyroid as a result of many different disruptions in the further metabolism of iodide in the thyroid gland.
Acknowledgments
I am most grateful to the following for careful criticisms of the manuscript: Dr. Jacob Robbins, NIH; Dr. John B. Stanbury, Boston; and Dr. Wilmar M. Wiersinga, Amsterdam.
Note: Since submission of this paper, an outstanding and extensive review on amiodarone and the thyroid has been published by W. M. Wiersinga (in Handbook of Experimental Pharmacology 128:225–287, 1997), which the reader should consult for additional information. In addition, interesting findings regarding anion selectivities have been published for the symporter expressed inXenopus oocytes (Eskandari, S., Loo, D. D. F., Dai, G., Levy, O., Wright, E. M., and Carrasco, N.: J Biol Chem272:27230–27238, 1997). These selectivities differ from nearly all previously published material in that perchlorate, and the closely related perrhenate, ions are not transported, whereas accumulation in thyroid tissue has been repeatedly demonstrated. By contrast, anions weakly active in the thyroid, such as chlorate and nitrate, were well transported in the oocyte system. These discrepancies may be due to differences in the number of compartments; differences in the metabolism of some of the anions; the high anion concentrations used; or the requirement for an additional factor for accumulation. It will be instructive to resolve these differences.
Footnotes
-
↵FNa Address for correspondence: J. Wolff, Building B, Room 2A23, National Institute of Health, Bethesda, MD 20892.
- Abbreviations:
- AIT
- Amiodarone-induced thyrotoxicosis
- Amiodarone
- 2-butyl-3-[3,5-diiodo-4-(β-diethylaminoethoxy)-benzoyl]benzofuran
- ATPase
- adenosine triphosphatase
- TPO
- thyroid peroxidase
- TSH
- thyroid-stimulating hormone
- The American Society for Pharmacology and Experimental Therapeutics
References
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
In this issue




{kind=link}
{kind=link}
{kind=link}
Jump to section
- Article
- I. Introduction
- II. The Anion Transport System
- III. Chemistry, Metabolism, and Distribution of Perchlorate
- IV. Transport Effects of Perchlorate
- V. Anomalous Antigoitrogenic Effect of Perchlorate
- VI. The Clinical Use of Perchlorate
- VII. Amiodarone
- VIII. The Perchlorate Discharge Test
- IX. Summary
- Acknowledgments
- Footnotes
- References
- Figures & Data
- Info & Metrics
- eLetters